糖心vlog 刘西莉教授课题组成功揭示了辣椒疫霉效应蛋白PcAvh337b靶向拟南芥 AtMBP-1-Nats模块,进而促进病原菌侵染致病的分子机制。该研究首次证实,AtMBP-1蛋白在植物体内真实存在,且可作为感病因子参与植物免疫调控;同时进一步明确了AtMBP-1的蛋白稳态受到两种不同N端乙酰转移酶NatA与NatC的拮抗性调控。这一拮抗性的调控维持了AtMBP-1在植物体内的动态平衡及相对较低的蛋白水平。相关成果的发表,破解了学界长期以来围绕AtMBP-1蛋白能否在植物体内稳定存在的争议难题。
辣椒疫霉是一种极具破坏性的植物病原卵菌,可以侵染茄科、葫芦科等70多种植物,给农林业生产造成严重的经济损失。RxLR类效应蛋白作为辣椒疫霉重要的致病因子,其变异性极强,功能高度分化。解析该类效应蛋白的变异机制及其在病原菌侵染寄主植物中的生物学功能,可为植物免疫相关蛋白的挖掘和作物疫病的科学防控提供理论支持。刘西莉教授课题组在前期研究基础上,开展了效应蛋白PcAvh337变异及其调控植物免疫的机制研究。相关成果以“Phytophthora effector exploits WY module truncation to manipulate Nt-acetylation-mediated AtMBP-1 turnover and suppress plant immunity”为题,近期发表在Science Advances期刊。该研究揭示了病原菌通过截断PcAvh337的WY模块以促进效应蛋白的变异和功能分化,并解析了PcAvh337b通过特异性靶向AtMBP-1-Nats分子模块,打破了植物体内NatA和NatC介导的N端乙酰化修饰所维持的AtMBP-1动态平衡及相对较低的蛋白水平,进而抑制植物免疫反应并促进辣椒疫霉侵染的分子机制。
该研究通过转录组数据分析,筛选获得两个具有不同WY模块数量的旁系同源RxLR效应蛋白PcAvh337a&b。基因组数据库检索比对发现,PcAvh337b的C端存在TNT1-94转座子元件插入,导致其编码蛋白提前终止,造成第3个WY模块缺失,进而形成截断的PcAvh337b。功能研究表明,PcAvh337a&b均是病原菌侵染的关键毒性因子,且功能非冗余(图1)。
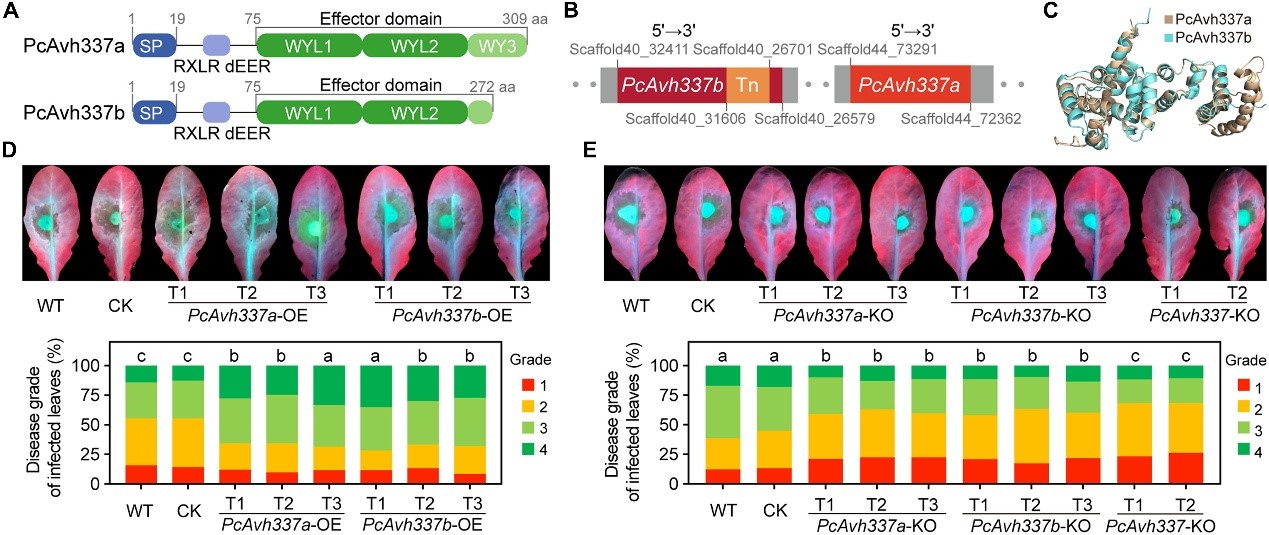
图1 PcAvh337a和PcAvh337b均是辣椒疫霉侵染的关键效应因子
通过寄主靶标筛选鉴定到PcAvh337b特异性与植物AtMBP-1蛋白互作。AtMBP-1被报道通过LOS2选择性翻译产生,并参与ABA信号通路的调控(Kang et al., 2013);也有研究指出,AtMBP-1在体内很难被检测到,可能并不能通过LOS2选择性翻译产生(Yang et al., 2022)。关于这一争议,该研究通过PCR扩增和IP-MS分析证实了AtMBP-1在植物体内真实存在(图2)。同时也首次发现,AtMBP-1参与植物免疫调控,其作为感病因子,可以抑制植物免疫相关基因的表达。进而明确了AtMBP-1蛋白稳态受到两个不同的N端乙酰转移酶NatA和NatC的拮抗性调控机制:NatA介导的N端乙酰化修饰增强AtMBP-1的稳定性,而NatC介导的N端乙酰化修饰促进AtMBP-1的降解。这一拮抗性的调控维持了AtMBP-1在植物体内的动态平衡及相对较低的蛋白水平。
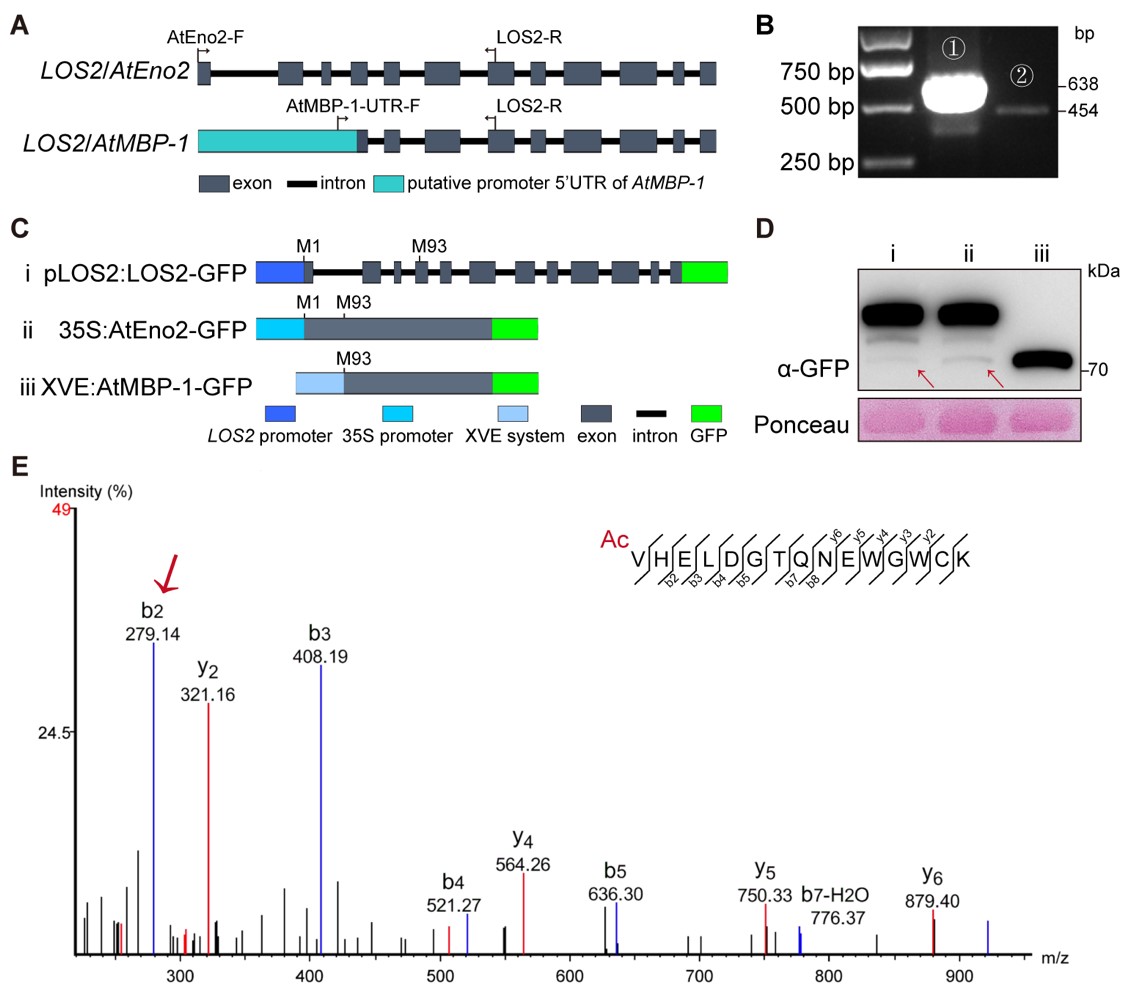
图2 AtMBP-1的转录和翻译分析
而效应蛋白PcAvh337b能够特异性靶向AtMBP-1-Nats复合物,打破NatA和NatC介导的N端乙酰化修饰所维持的AtMBP-1动态平衡,并通过促进AtMBP-1与NatA的结合,增强NatA介导的AtMBP-1的N端乙酰化修饰和降低AtMBP-1的泛素化修饰水平,以稳定感病因子AtMBP-1,进而抑制植物免疫反应并促进辣椒疫霉的侵染。
该研究揭示了病原菌效应蛋白变异和扩张的具体途径,解析了效应蛋白通过劫持植物感病因子AtMBP-1的N端乙酰化与泛素化修饰的交互调控系统,精准调控AtMBP-1蛋白稳定性,从而抑制植物免疫的分子机制。研究结果为深入理解病原菌与寄主植物的互作机制提供了新的视角,也为后续作物抗病遗传改良提供了潜在的基因资源和理论支持。
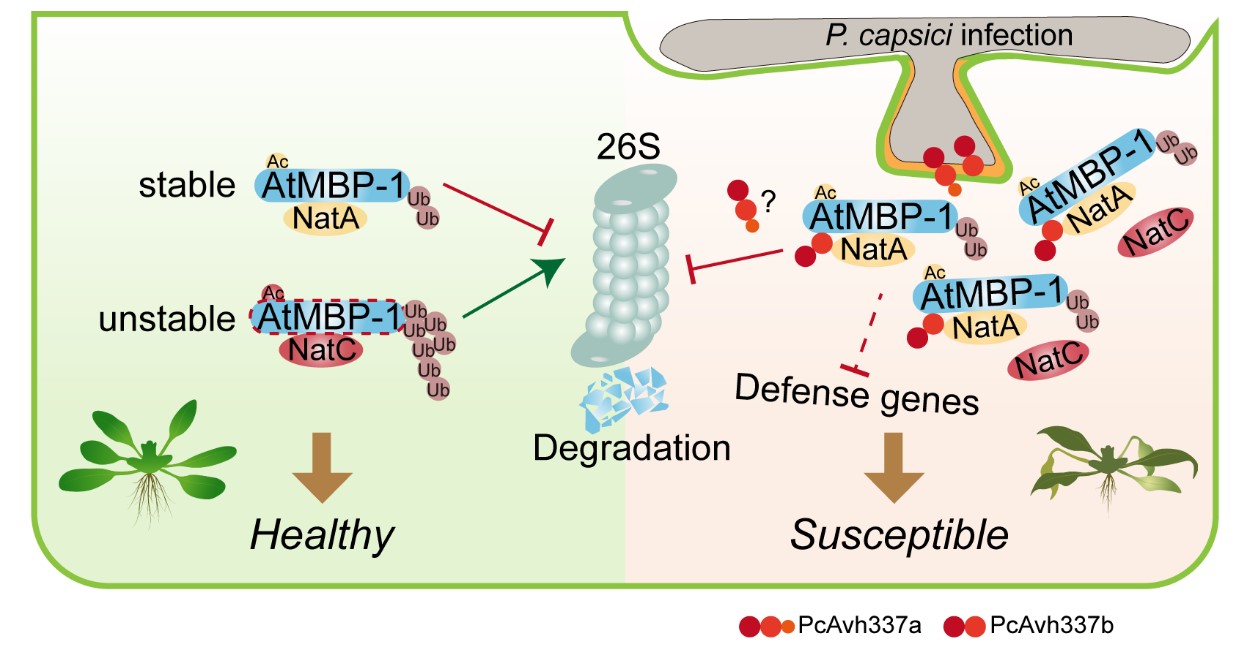
图3 PcAvh337b调控AtMBP-1-Nats模块以稳定感病因子AtMBP-1和促进辣椒疫霉侵染的模式图
糖心vlog 已毕业博士研究生田嵩为论文的第一作者,刘西莉教授和顾彪副研究员为共同通讯作者。该研究得到了国家自然科学基金(32270211)、陕西省自然科学基金(2022JM-118)和陕西省创新能力支撑计划(2020TD-035)的资助。
原文链接://www.science.org/doi/10.1126/sciadv.ady1482
编辑:刘小凤
审核:赵 磊
